
2000年2月 No.9

1999年7月 アラスカフェアバンクス郊外北方森林での人工森林燃焼実験
FROSTFIRE共同研究として参加。(福田正己)
写真の出展: http://www.fsl.orst.edu/fera/frostfire.html
低温科学研究所所長 本堂 武夫
20世紀を総括する新聞記事やテレビ番組を目にすることの多い昨今であるが、科学の世界も例外ではない。ある科学雑誌に「20世紀に起きた主要な発見・発明とその関連図」というのがあった。図の中央に、物理学、化学、生物学・医学の3本の太い流れがあって、その両脇に宇宙科学と地球科学の細い流れがある、ざっとそんな図柄である。宇宙科学と地球科学は、世紀を通じてほぼ同じ細い流れで続いているが、中央の3つの主流に大きな変化が見て取れる。流れの太いところは、同時代に多くの発見・発明があったことを物語っている。20世紀初頭は、化学の流れが太くなっており19世紀の華やかさを引き継いでいる。1930年頃、物理学の流れがもっとも太くなり、世紀後半は生物学・医学太くなっている。
このような流れの変化にはきっかけがある。1930年前後の物理学の高揚は、量子力学の登場によるものであり、世紀後半における生物学の隆盛は、DNAの構造が解明されたことを契機とするものである。量子力学は、ニュートン以来の常識を覆して、それまでの自然観を一変させたという点で革命的であり、DNAの解明は、生命の本質に迫る新たな一歩を踏み出したという点で画期的なできごとであった。
世紀後半では、物理学も化学も流れが細くなっているが、これは決して衰退を意味しているのではない。これらの学問が、広範な学問領域に広がっていったために、境界がはっきりしなくなったというべきであろう。量子力学は、歴史的評価に耐え得る大発見だったからこそ、その後広範な学問に影響を与え続け、次々と新たな発見を生み、ミクロの世界を支配する標準的な法則として自然科学の基礎になった。同時に、物理学と化学のオーバーラップは広い範囲にわたるようになったし、両者と生物学との境界もまたしかりである。
20世紀も終わり近くになって、コンピュータの急速な進化が科学の世界に大きな影響を及ぼした。分子動力学法のような新たな手法が生まれたことももちろん重要なできごとであるが、なによりも、コンピュータは研究室の風景を変えてしまった。実験装置の多くはコンピュータ化され、最適化された測定や高度なデータ解析が、専門的な知識なしで、できてしまう。かっては、解析不可能であった多体問題や非線形問題の解答をいとも簡単に与えてくれる。計算手法や実験装置の隅々まで知り尽くさないと納得できない旧世代型の研究者にとって、なんとも不可解な時代になってしまった。
しかし、見方を変えると、このような研究スタイルの変化は自然な成り行きであるように思える。生物の固体発生が、進化の系統発生を時間短縮して繰り返すように、研究が進化するためには、それまでの知の資産を凝縮する道具として、コンピュータが不可欠な存在なのだ。現代の科学は、実に広範な知識の蓄積の上に成り立っており、もはや、一人の研究者が、関連する様々な理論や手法をすべて身につけることは不可能である。広範な分野の成果を取り込むには、それぞれの専門家の協力が不可欠であるが、コンピュータはこれを一人でもできるようにしたと言えよう。高度なデータ解析の手法は、数理科学の成果であるし、結晶構造解析の自動化は、結晶学の成果である。それらの専門分野の長年にわたる成果が、ソフトウエアや装置に凝縮されている。いわば、専門家の手助けを間接的に受けているようなものである。もちろん、装置や手法を隅々まで知ろうとする姿勢は、研究者の基本である。たとえ旧世代型と言われようと、この点にはこだわりたいものである。しかし、それぞれの分野の成果が、汎用的な装置やソフトウエアという形で開放されることによって、その応用範囲は飛躍的に拡大し、まったく新たな展開をみせるのも事実である。
一方、読売新聞が昨年末に出した特集「20世紀の主役たち」を見ると、自然科学の分野に中谷宇吉郎の名前がある。もちろん、雪の研究が認められてのことである。人工雪の研究は、1930年代に行なわれており、上記の図で物理学の流れがもっとも太い時期と重なっている。雪の研究は、量子力学とはなんの関係もないが、この時期にこのようなユニークな研究が生まれたということに興味を覚える。雪の研究は、身近な自然現象の典型的な例であるのに対して、量子力学は日常体験からはもっとも遠いところにあって、不確定性原理に見られるように、その解釈は人々の常識をはるかに超えている。量子力学が物質の究極の姿を求めた結果であるのに対して、雪の研究は自然のあるがままの姿を描く試みであったと言えるかもしれない。人工雪の装置は、驚くほどシンプルである。しかし、この単純さの中に、今日で言う複雑系の科学に通ずるものが秘められている。
複雑系は流行語になってしまったが、量子力学とは違った意味で、革命的である。ニュートン力学に従う巨視的な系でも未来予測不可能な場合がある。わずかな初期条件の違いが、まったく違う結果をもたらす場合があるかと思えば、複雑な自然現象が意外に単純な法則をもっていたり、これまでにない科学の新しい側面が見えてきている。複雑な現象を要素に分解して、純粋化した過程について美しい法則を求めてきた要素還元型の研究に限界がみえている。自然の階層構造にあって、ミクロ世界を支配する法則がすべての階層を記述するのに適しているとは限らない。それぞれの階層を支配する法則があるはずである。次の時代に開花するのは、そういう研究かもしれない。巨視的な系、言い換えると身近な系、の科学が、コンピュータの進化とあいまって、飛躍的に進歩する可能性はおおいにあろう。
奇しくも、西暦2000年は、中谷宇吉郎生誕100年にあたる。そんな年のはじめに、大学や研究所をめぐる昨今の複雑怪奇な状況を離れて、科学に夢を託したいと思った次第である。
福田正己(寒冷圏総合科学部門)
1997年12月の温暖化抑制のための京都国際会議(COP3)では、二酸化炭素などの温暖化効果ガスの排出規制について、具体的な対策構築が話し合われた。1995年のIPCCによる報告では、年間55億トンもの二酸化炭素が、化石燃料燃焼により大気へ放出されている。放出と吸収の収支か、約7.7億トンが北方森林へ吸収されていると推定された。これがCOP3の議論に影響を与え、アメリカ・ロシアなどが、森林の吸収分を加算した排出規制案(ネット方式)を提案した。しかし、果たして北方森林は二酸化炭素の吸収源となっているのか、多くの疑問が残された。1997年にFAOが公表した1964年−1994年の30年間世界の森林の森林面積増減では、熱帯南米(アマゾン)の約10倍ものシベリアの森林が消失していた。その主な原因は、森林火災と伐採であり、人為起源の原因であった。
更に、1991年来のシベリア永久凍土調査から、天然ガスパイプラインからの漏洩が、メタンガスの供給源となっていることも予想された。こうした温暖化ガス排出規制の鍵を握るのは、永久凍土地域での人間活動ではないかと考えるにいたった。そこで、本研究プロジェクトでは、温暖化ガス発生の要因となる人為による永久凍土攪乱とそれが将来の温暖化に及ぼす影響を評価することを目的としている。また、新たな地球環境課題への取り組みとして、原因究明→予測→抑制までの一貫した流れでの課題達成を掲げた。そのためには、地球物理学、森林科学、大気化学といった基礎科学に加えて、燃焼工学、システム工学といった、応用工学分野に跨る学際的な研究体制をとることとなった。
永久凍土攪乱はどのようにして起こるか。その最大の原因は森林火災である。そこで森林火災による二酸化炭素収支変動を明らかにするために図1に示すような、森林火災による変動を現地で観測する方法をとることとなり、シベリアヤクーツク近くのタイガ林内に観測タワーを設置し、長期の熱−水−炭素収支のモニタリングを行っている。(写真1)。森林火災は直接的に大気へ二酸化炭素を放出するだけでなく、焼失による森林の二酸化炭素吸収機能の低下、熱収支バランスも乱れに起因する永久凍土の融解とその過程で貯留されていたメタンガスの放出という複合した過程で温暖化効果ガスの放出源となる。それを正確に評価し、また抑制法を確立する。そのための研究手法を模式的に図1に示す。また、シベリアに敷設された天然ガスパイプラインが、その不十分な対策から凍土融解や凍上のため破損し、そこからメタンガスが大量に放出されている。そこで永久凍土地域でのガスパイプラインの敷設に伴う、技術的問題解決も重要課題である。アラスカフェアバンクス郊外の永久凍土地域では、実際にパイプラインを地下埋設し、凍土融解や凍上によるパイプ挙動の現地観測を行う大規模実験を行っている。その実施サイトの平面図と敷設工事の様子を写真2に示す。
本研究プロジェクトは、北海道大学をはじめとする大学、国立研究所、民間企業の研究者合計22名から成り立っている。
なお本研究プロジェクトについては、以下のホームページが開設されており、具体的なメンバー構成も公表されている。
http://frost2.lowtem.hokudai.ac.jp/pdgg/
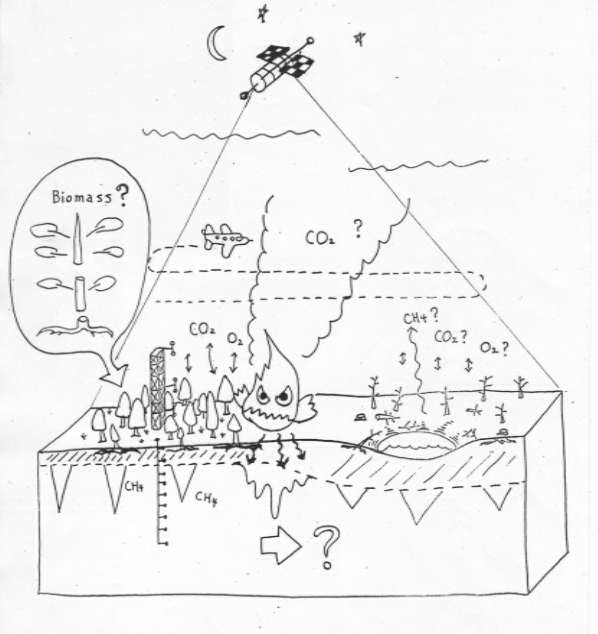
図1 森林火災が温暖化ガスに与える影響

写真1 シベリアタイガにおける熱−水−炭素収支観測東シベリアヤク−ツク周 辺ネレゲル地区

写真2 ガスパイプラインの実スケ−ル実験 アラスカフェアバンクス実験地パイ プの施設 1999年12月
芦田 正明(低温基礎科学部門)
研究のおもしろさは時として考えもおよばなかった方向に発展する場合があることである。昆虫血液が試験管の中に取り出されるとなぜ黒色色素(メラニン)を合成し始めるのかについての研究もその一例である。メラニン合成の背後に隠された仕組みは昆虫のカビやバクテリアに対する主要な生体防御機構の一つであることが1990年代に入りはっきりしてきた。此の機構で使われている分子と相同な分子が我々人間を含めたほ乳類の自然免疫に関わっているらしいことも最近明らかになった。さらに、我々の研究が基となって家蚕(カイコ)の血液がグラム陽性菌とカビの検出試薬として市場に登場し、大海原におけるバクテリアの存在量を測る手だてとして利用される例などが知られるようになった。研究の背景と最近の話題を紹介する。
メラニンはチロシンやドーパなどのフェノール性物質が空気中の酸素の存在下で酸化され、生じたキノン類が重合した複雑な化合物ある。フェノール性物質の酸化反応を触媒するのがフェノール酸化酵素である。昆虫血液にはチロシンやドーパが高濃度で存在している。フェノール酸化酵素も血液中に存在している。血液には空気中の酸素が充分溶け込んでいるにもかかわらず、なぜin vivo(体内)でメラニンは合成されないのだろか。半世紀近く以前に私の恩師である大西英爾博士(現名古屋大学名誉教授)がフェノール酸化酵素は不活性な前駆体(フェノール酸化酵素前駆体)として昆虫血液に存在していることを発見された。1951年のことである。この発見は、血液が体外に取り出されるとフェノール酸化酵素前駆体の活性化反応が始まることを明らかにした。このときから、次の2つの疑問 ”活性化の際、前駆体分子にどの様な変化が起こるか”と”活性化反応の開始はどの様に制御されているか”に答えるための長い研究の歴史がはじまった。まず、フェノール酸化酵素前駆体とそれを活性化する因子(フェノール酸化酵素前駆体活性化酵素、PPAEと以下略称)がカイコ幼虫から精製され、試験管の中で活性化反応が解析された。フェノール酸化酵素前駆体はPPAEの作用で分子中のたった一つのペプチド結合が加水分解されることによって活性型に変換されることが明らかになった。加水分解されるのはフェノール酸化酵素前駆体分子のアミノ末端から数えて50番目のアミノ酸(アルギニン)のカルボキシル基がわのペプチド結合で、−アスパラギン−アルギニン−フェニルアラニン−グリシン−というアミノ酸配列の中にある。このアミノ酸配列はショウジョウバエやタバコスズメ蛾のフェノール酸化酵素前駆体にも存在している。ちょっと余談になるが、ここでフェノール酸化酵素の一次構造(アミノ酸の配列順序)についての我々の研究から明らかになった興味ある事実にふれておきたい。昆虫のフェノール酸化酵素は節足動物のヘモシアニン(ほ乳動物の血液中に存在するヘモグロビンのように酸素を運搬する機能を持つ銅タンパク)と相同なタンパクで脊椎動物、植物、細菌のフェノール酸化酵素とは進化の上で起源の異なる分子であることが判明したことである。下等節足動物はほとんど皆ヘモシアニンを持っているにもかかわらず、昆虫はこれを持っていない。昆虫は空気中の酸素を直接組織に運ぶ気管系を発達させたので、ヘモシアニンを必要としなくなりこの分子を失ってしまったのだと考えられてきた。昆虫は必要でなくなったヘモシアニンをフェノール酸化酵素前駆体として廃品利用されたのかも知れない。
フェノール酸化酵素前駆体がPPAEによる限定加水分解で活性化されることははっきりした。それではPPAE活性はどの様に制御されているのだろうか。昆虫の特殊な生体防御機構の研究からこの問いにたいする答えが得られることになった。
昆虫の生体防御機構として、血球細胞による細菌の貪食や大型異物を取り囲む被嚢形成、血液凝固、抗菌ペプチドの合成などが知られている。これらの機構は昆虫のみならず脊髄動物や他の無脊椎動物にも広く分布していることが知られている。生まれつき備わっている機構なので自然免疫とよばれている。高等脊椎動物にはこれ以外に免疫グロブリンなどが関与する獲得免疫の機構があるが、自然免疫の仕組みは生物進化の課程で獲得免疫よりずっと前に出現したと考えられている。植物のカビやバクテリアにたいする生体防御機構に動物の自然免疫に使われている分子と相同な分子が組み込まれていることもごく最近発見されているいる。
昆虫や一部の下等節足動物では体内に侵入したカビの菌糸やバクテリアの周囲、傷害部位、宿主選択を誤って生みつけられた寄生蜂の卵の周囲にメラニンが沈着することが昔からしばしば観察されてきた。この異物周囲でのメラニン合成は昆虫病理の研究者の間で昆虫の生体防御反応のおおきな特徴として注目を集めてきた。フェノール性物質からメラニンが合成される過程で生じるキノン類は反応性が高く、異物周囲で生じる不溶性高分子物質をキノンなめしにより物理的に強固にして異物を封じ込めてしまうからである。また、キノンには細胞毒性があり細菌類を殺すのにも役だっている可能性が指摘されている。
カビやバクテリアの周囲でメラニンが合成されるということは、そこでフェノール酸化酵素が働いていることを意味している。フェノール酸化酵素はフェノール酸化酵素前駆体がPPAEにより活性化されて生じる。このことから考えるとPPAEの活性制御にこれら微生物が関係していそうである。どの様に関係しているかを調べるには昆虫血液をin vivoの状態を保ったまま試験管の中に移しとることが是非必要である。なぜならば、前にも述べたように血液を体表に傷をつけて採取したのでは、得られた血液の中でPPAEは働きだしてしまうからである。”in vivoの状態を保ったまま”ということが難しく1980年代はじめまで世界中の誰もカビやバクテリアとフェノール酸化酵素前駆体の活性制御を結びつけることはできなかった。1980年に筆者はカイコの血液(普通は黄色をしている)を様々な(他の研究者がちょっとやらないような)方法で採取することを試みていた。もちろん”in vivoの状態を保ったまま”の血液を得るためである。紙面の都合で詳細を記すことができないが、とにかく試験管のなかでいつまでも黄色で、カビの細胞壁の成分であるザイモサン(βー1、3ーグルカンとマンナンの混合物)を加えると黒化する血液を得ることに成功した。この血液の血漿成分のおかげでカイコ血液におけるメラニン生成機構(フェノール酸化酵素前駆体活性化機構)の研究が初めて可能になった。
”in vivoの状態を保ったまま”得られた血液を用いて明らかになったフェノール酸化酵素前駆体活性化機構を図1に示す。予想されたようにフェノール酸化酵素前駆体はカビやバクテリアの細胞壁成分により活性化されることが証明された。しかも、フェノール酸化酵素前駆体はバクテリアとカビの細胞壁成分により直接活性化されるのではなく、バクテリアとカビの細胞壁成分であるペプチドグリカンやβー1、3ーグルカンと特異的に結合する認識タンパク(ペプチドグリカン認識タンパクとβー1、3ーグルカン認識タンパク)、いくつかのセリン型プロテアーゼ(活性中心にアミノ酸のセリンを持つタンパク分解酵素)前駆体などから構成されるカスケードの作用で活性化されることが明らかになった。ペプチドグリカン認識タンパクとβー1、3ーグルカン認識タンパクがそれぞれのリガンド(ペプチドグリカンとβー1、3ーグルカン)に結合することによってカスケードの引き金が引かれる。 このカスケードは世界中の研究者によりプロフェノールオキシダーゼカスケード(略称ではproPOカスケード)とよばれるようになった。このカスケードは昆虫の異物認識機構の一部を構成し、昆虫の主要な生体防御機構の一つとして認められている。ProPOカスケードの構成要素はまだ図1に示したものしか単離されていない。どのくらい多くの構成要素が存在するのか、私たちの研究でおおよその見当はついているが、それらをすべて精製するにはまだまだ時間がかかりそうである。精製されたすべての構成要素を用いてin vitroでカスケードの活性化反応が解析できるようになればカスケードの生理的役割に関する我々の知識はおおいにいに広がるだろう。我々は主要な努力をこの方向に向けて研究を進めている。
ペプチドグリカン認識タンパク(peptidoglycan recognition protein,PGRP)は吉田英哉博士(現、岡山大学・資源生物研究所)が私の研究室にいる時に精製し、ごく最近、落合正則博士がその一次構造を明らかにした。驚いたことに、PGRPと相同なタンパクが人間やマウスにも存在し、機能が判らないタンパクとして放置されていたことである。バクテリアの細胞壁成分であるペプチドグリカン(PG)はほ乳動物でも非自己として認識され発熱など様々な生理作用を示すことが知られているが、PGに特異的に結合する分子はほ乳動物では知られていなかった。我々のPGRPについての研究が発端となってほ乳動物でPGが非自己として認識される仕組みや、生理活性を示す仕組みについての研究が急速に進展することが期待されている。さらに、proPOカスケードがカビの細胞壁成分の * ー1、3ーグルカンにより活性化されて生じるプロテアーゼ(図1でBAEEaseとして記されている酵素)は脂肪体(昆虫でほ乳類の肝臓のような働きをしている組織)の細胞にカビを殺す作用のある小さなタンパクを合成するように指令するのに働いているらしいことも判ってきた。proPOカスケードはメラニンを合成するためのみに働くのではなく、カビやバクテリアを非自己として認識し、その認識シグナルを増幅して、昆虫の他の生体防御機構が働き始めるためのシグナルを提供する仕組みだったのである。
カブトガニの血液凝固系はセリン型プロテアーゼ前駆体などから構成されるカスケードで、グラム陰性菌の細胞壁成分であるのリポポリサッカライド(lipopolysaccharide,LPS)やカビの細胞壁成分であるβー1、3ーグルカンにより引き金が引かれるカスケードである。このカスケードは医療機器や医薬のグラム陰性菌あるいはカビによる汚染を検査するテスト、”リムルステスト”(リムルスはカブトガニの属名)として広く使用されている。リムルステストでは1ml中に数ピコグラムのLPSやβー1、3ーグルカンあればそれらを検出できる。リムルステストはこのように感度の高い優れた汚染検査試薬であり、現代の医療、医学・生物学研究には欠くことのできない試薬となっている。多くの先進国では医療機器や医薬のリムルステストによる汚染検査が義務ずけられている。しかし、このリムルステストに泣き所がある。1つは、グラム陽性菌を検出できないことである。グラム陽性菌は多くの感染症のを引き起こすことが知られている。現在問題になっている院内感染病原菌、メリチシン耐性菌もグラム陽性菌である。さらに、カブトガニは世界的に資源が減少している。日本では天然記念物に指定されているほどである。リムルステストにたいする需要は益々増加することが予想されているので、将来需要をまかなうに充分なカブトガニ血液の供給が困難になるかもしれない。これらの事情がリムルステストに代わる感度の良い細菌汚染検査試薬、あるいはリムルステストの弱点を補完する試薬の登場が期待されている理由である。今から10年近くまえ、リムルステスト試薬を発売している薬品会社の1つである和光純薬工業の大阪研究所に勤務する土谷氏から電話を頂いた。カイコのproPOカスケードを汚染検査薬として利用する可能性を研究したいと言うお話だった。私たちの研究を基に、土谷氏らはカイコ血液のproPOカスケードを汚染検査薬として育て上げるために様々な工夫をされ、ペプチドグリカンとβー1、3ーグルカンを検出する試薬としてカイコ血液からSLP試薬を製品化することに成功された。4年前のことである。SLP試薬による汚染検査にはプレートリーダーのような比較的高価な機器を使用する方法と機器を使わず目視によるエンドポイント判定法がある。図2に目視によるエンドポイント判定法でペプチドグリカンとβー1、3ーグルカンを検出した例を示した。SLP試薬について今後解決しなければならない問題点は色々考えられるが、さしあたって次の2点が重要である。第一は、ペプチドグリカンあるいはβー1、3ーグルカンだけを検出するSLP試薬を開発すること(実験室段階ではそれぞれに特異的な試薬の調製が可能である)、第二はペプチドグリカンに対する試薬の感度をたかめることである。現在のSLP試薬のペプチドグリカンにたいする感度が低い原因の一端はペプチドグリカンの物理的性状にあると考えられている。ペプチドグリカンは可溶性の物質ではない。したがってSLP試薬に加えられたペプチドグリカンのすべてがproPOカスケードの引き金を引くのに有効に働いていないのかもしれない。
ペプチドグリカンは様々な生理活性を持つことが知られているが、ほ乳動物の体内での動態についての研究は良い検出法がなかったために不明な点が多かった。SLP試薬はグラム陽性菌による汚染を検出するたに初めて実用化された手段であるだけでなく、ペプチドグリカンの薬理作用研究の新しい展開を促す触媒として機能するかもしれないと期待されている。また、上に述べた問題点を克服することによってさらに使いやすい試薬として用途が広がり多くの人に使用されるようになるだろうと思われる。東京大学・海洋研究所の小暮研究室の柴田晃博士はSLP試薬を大海原のバクテリアのバイオマスを定量するのに応用しているとお聞きしている。これなどは予想だにしなかった分野でのproPOカスケードの利用例である。
昆虫血液が体の外に取り出されるとその色が黒くなる。その理由を調べてみると、メラニン合成はカビやバクテリアにたいする昆虫の生体防御機構が働いた結果だということが明らかになった。さらに、予想もしなかった方向に研究は展開し、グラム陽性菌を検出する試薬の開発に貢献することができた。ProPOカスケードはほ乳動物血液の補体系や血液凝固系に匹敵すると看なされ、現在では昆虫の生存にとって欠くべからざる重要な生体防御系だと考えられるようになった。昆虫のproPOカスケードに関して現在われわれが知っている重要なことがらのほとんどすべては我が研究室で明らかにされたと断言しても過言でない。これらの成果を得ることができた最大の原因は、昆虫の血液が黒くなる仕組みに興味を持ちそれをを持続して研究したことだと思っている。フェノール酸化酵素が前駆体として存在することが報告されてから半世紀が経過した。この先の研究で今まで以上に興味ある現象に遭遇できるのではないかと秘かに期待している。
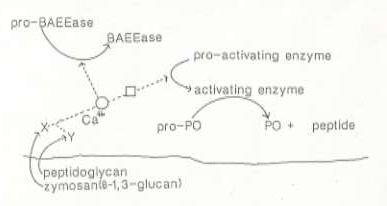
図1 カイコ血漿中のプロフェノールオキシダーゼカスケード。X,βー1、3ーグルカン認識タンパク;Y,ペプチドグリカン認識タンパク;BAEEase,ベンゾイルアルギニンエチルエステルを加水分解する酵素として発見されたセリン型プロテアーゼ。現在、生体内での機能に注目が集まっている;pro−BAEEase、BAEEaseの前駆体;PO,フェノール酸化酵素;pro−PO、フェノール酸化酵素前駆体;activating enzyme,pro−PO活性化酵素(セリン型プロテアーゼの1種。本文ではPPAEと略称されている);pro−activating enzyme,activating enzymeの前駆体。マル印はカルシウムイオンを必要とする反応を示す。ProーBAEEase とpro-acti- vating enzymeは限定加水分解により活性化されることが証明されている。図で点線はまだ解明されていない部分を示している。
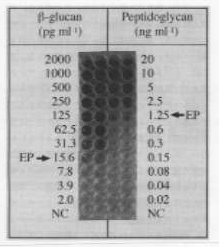
図2 SLP試薬による目視テスト。SLP試薬と同容量の試料を混合し30度1時間放置後、結果を判定した。βー1、3ーグルカンとペプチドグリカンのエンドポイント(EP)はそれぞれ15.6pg/mlと1.25ng/mlであることが示されている。
渡部直樹・香内 晃(低温基礎科学部門)
私たちはこれまでに宇宙空間(暗黒星雲,太陽系星雲など)に存在する氷微粒子の生成・変成機構やアモルファス氷の物性研究を通して,太陽系の氷天体の起源や進化を議論してきた.この種の研究は今後も引き続き行っていくが,本稿では現在進行中の新しい研究を紹介する.
はじめに,暗黒星雲での物質進化研究の重要性を述べる.暗黒星雲には鉱物,有機物,アモルファス氷からなる固体微粒子が存在し(図1),それらは太陽系を作るもとになった物質である.気相には,H,O,C,Nなどの原子が存在する.暗黒星雲は温度が10Kと低いにもかかわらず,固体微粒子表面では水素原子のトンネル反応等により活発な化学反応が進行し,種々の物質が形成される.これらの物質には気相反応では生成し得ない重要な分子種が数多く含まれており,アモルファス氷上の表面反応は星間分子,氷,有機物生成の鍵となっている(図1).したがって,暗黒星雲における物質進化の研究は暗黒星雲それ自体の研究だけでなく,太陽系や生命の起源を明らかにする上でも重要な意義を持っている.また,極低温アモルファス氷上での表面反応は物理・化学的にもほとんど手が付けられていない興味深い研究対象である.しかし,これまでの研究では,これらの表面反応はほとんど無視されてきた.また,紫外線照射による光化学反応も物質の生成・変成を考える上で重要になるが,定量的な研究は全くなかった.
そこで,私たちは,暗黒星雲で起こりうる現象,特に,表面反応と光化学反応を実験室で忠実に再現し,星間分子,特に,水素分子,アモルファス氷,有機物の生成・進化過程を定量的(生成反応経路の決定,反応速度,反応断面積の測定,生成後の分子の内部状態等の解明)に理解しようとしている.
私たちは図2のような真空装置を用い,アモルファス氷薄膜を作製し,様々な分子種に関して表面反応による生成メカニズムを調べている.この際に問題になるのが,H,O,Nなどの原子源である.通常,H2などの分子に何らかの方法でエネルギーを与えて分解し,原子を作る方法が用いられている.ところが,通常の原子源には原子の他に,分子,イオン,ラジカルなどの不純物が含まれており,我々の研究目的には使用できない.不純物の全く含まれない基底状態の原子だけからなる原子源を,いかに開発するかが研究の鍵となる.5〜6年をかけて種々の原子源を試作・評価してきたが,最近ようやく実験に用いることのできる原子源が完成した.また,氷表面で生成した水素分子,アモルファス氷,有機物などをどのような方法で検出するかも大きな問題であった.こちらは,超高感度表面赤外線分光で検出するめどがたった.氷表面上に吸着した1/10〜1/100層の厚さの水素分子の赤外線吸収スペクトルが測定できるようになった.
アモルファスH2O氷への紫外線照射による水素分子の生成反応,アモルファスH2O-CO氷への紫外線照射によるCO2分子の生成反応は,定量的なデータの取得(生成反応経路の決定,反応速度,反応断面積の測定)に成功した.今後は,不純物を含むアモルファスH2O氷への紫外線照射による有機分子の生成反応などの紫外線照射反応の定量化を行うとともに,原子の関与した表面反応の定量的実験を推進していきたい.このようなデータが得られてはじめて,暗黒星雲での物質(化学)進化の実体が明らかになるであろう.
超高感度表面赤外線分光装置の作製にあたって,渡部に交付された所長リーダーシップ経費を使わせて頂いた.ここに記して感謝致します.
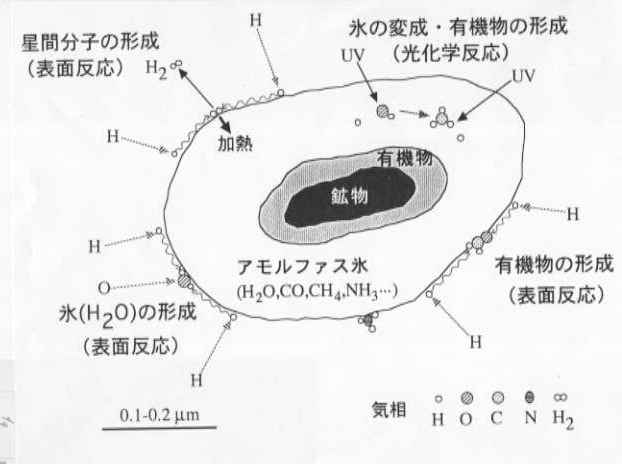
図1 アモルファス氷星間塵上での物質生成

図2 超高真空・極低温表面化学反応実験装置
竹内 謙介(寒冷海洋圏科学部門)
低温研に来た頃、私はエルニーニョの研究をしていました。エルニーニョは熱帯の現象で低温研とはあまり縁がなさそうですが、低温研に来る気になったのは気候変動にとって熱帯に劣らず、高緯度海洋が重要であると考えたからです。
エルニーニョは大気海洋相互作用で起きることが明らかになった最初の気候変動現象です。エルニーニョに対する関心が高いのは、そのインパクトが大きいからですが、その理由は、大気の循環のエンジンにあたる熱帯でおきるからです。大気は加熱されると上昇し対流を起こしますが、加熱は熱帯で最も大きいので変動は広く行き渡りやすいことになります。大気と海洋のシステムを二つの流体が重なったものと考えると、対流に関して、海面をはさんで上下逆の関係になっていて、大気とは逆に海水が冷やされると沈降し対流を起こします。つまり、大気の熱帯にあたるのが海洋の高緯度にあたると言えます。
もうひとつ海氷の存在も重要です。海氷は実にいろいろな役割をしています。大気と海洋の熱のやりとりを遮断する効果、太陽放射を反射する効果、塩分の再分配効果、等です。逆に、海氷は気候変動の影響を非常に受けやすいので、海氷のある高緯度海洋が気候変動で重要な働きをしているのは当然予想される事です。北海道の北にあるオホーツク海は世界的に見ても大規模な海氷が発達する海域としては最も低緯度にあります。また、オホーツクの海氷は年々変動が非常に大きいことでも知られています。この様なオホーツク海がすぐそこにある、と言うことは我々にとって有難いことです。ということで、私の研究室ではオホーツク海での大気海洋相互作用を中心に研究していますが、その中のいくつかを紹介します。
冬季、季節風の噴出しのときには、オホーツク海上では海面水温と海上気温との差が数十度にも達し、海氷がなければ大気海洋間の熱フラックスは非常に大きくなりますが、海氷があればかなり減ると考えられます。オホーツクでの熱フラックスの大きと仕事の差は大気の循環にも影響を与えることが考えられます。学生の本田さん(現地球フロンティア)が立花さん(現東海大学)とこの問題を扱いました。海氷の多い年と少ない年をいくつか選び出してそれぞれの年の冬の気圧場を平均したところ、オホーツクで気温が高く、低気圧になることは想像どおりでしたが、さらにベーリング海に高気圧、その東に低気圧と高低気圧の連なるパターンが現れ、アラスカでも7度も気温が上昇することが見つかりました。
しかしどちらが原因でどちらが結果なのか、データ解析だけで判断することは困難です。そこで地球環境の山崎教授の協力を得て、大気大循環モデルを用いました。オホーツクの海氷が多い場合と少ない場合を、そのほかの条件は同じにして計算してみると、データ解析の結果と良く似た高低気圧の列が再現されました。また、東大の山本さんの協力による解析で、このパターンがオホーツクの熱フラックスの変動の影響が偏西風下流方向にロスビー波という波として伝わっているものであることがわかりました。これまで、大気の変動による海氷への影響という研究はいくつかあるのですが、その逆例としては貴重なものです。
そもそも比較的緯度の低いオホーツクで海氷が発達する理由として、シベリヤの強い寒気の発達ととともに、オホーツク特有の海洋構造が挙げられてきました。アムール川からの淡水の流入によって海面に塩分の薄い層ができ、冷却時に深い対流が妨げられて凍りやすい、と言うものです。これが本当なら、アムール川の流量の変動と海氷面積の変動に関連があるはずです。大学院生の小木さんが、道教育大釧路分校西尾教授の下で立花さんと研究をはじめ、現在も継続している研究によると、不思議な事にアムール川の流量が多い年には海氷面積が小さいという、想像とは逆の相関が見つかったのです。
これに関しては幾つかの原因が考えられます。一つは両方とも最近注目されている10年スケールの気候変動の結果であるというもの。もう一つは、アムール川から流入する水が結構温度が高く、海水温を上げている可能性です。確かに秋の海面水温はアムールの河口付近を中心に、アムールの流量との相関が高くなっています。オホーツクの海氷面積に関しては地球環境の山崎教授や海洋動態研究室の木村さんの研究によると風の西風成分の強さが関係しているようです。つまりサハリンに沿って南下している海氷が西風で東に広げられると面積が大きくなると言う物です。しかし、それだけでは説明がつかない部分があり、それがアムール川の流量である可能性があります。
先ほどの数値モデルの時には海面は全く海氷に覆われて隙間がないと仮定しましたが、実際にはかなり割れ目や隙間があります。そのような隙間からの熱フラックスはかなり大きいと思われます。一つ一つの割れ目で計った例はあるのですが、必ずしも単独の効果ではない可能性があり、そのような隙間も含んだ海氷域全体でどうなっているのかを知るのは気候モデル作成のためにも必要です。そこでそれを調べるプロジェクトをここ3年ほどやっています。
アイデアとしては非常に単純で、季節風の噴出しの時、上流と下流で気温や水蒸気量の鉛直分布を気象ゾンデで調べれば、その空気がどれだけ海から熱や水蒸気をもらったかがわかるのではないか、と言う物です。上流としてはサハリンのユジノサハリンスク等、下流としては斜里を選び、また、毎年海上保安庁の巡視船宗谷に便乗させてもらい観測をしています。サハリンでは通常、1日1,2回のゾンデ観測を期間中2,4回に増やしてもらっています。斜里では泊り込みで、今年は1ヶ月半ほど、やはり一日2,4回のゾンデ観測をしています。この観測では数多くの教官や学生の人に協力してもらっています。やってみるとなかなかちょうど良い風が吹く事が少なく、また、海氷の量は完全に自然任せですから思うようにならないのですが、今年が最後の観測になるのでなんとか結果を出したいとおもっています。

隅田 明洋(寒冷陸域科学部門)
樹木の幹・枝は、光合成に必要な水や養分を通すパイプ役を果たすと同時に、光合成器官(葉)を機械的に支持してその空間分布を決める。幹や枝を細く長くすれば隣接する樹木より早く高くなってより多くの光を受けることができるが、光合成生産に必要な水を十分供給することはできないし、葉を支えるための力学的強度も低くなる。樹木の形状・構造には、それによって生じる利益と不利益の間に様々なトレードオフの関係が生じる。
常緑広葉樹アラカシ(Quercus glauca)の幹の成長パターンを調べてみると、光が十分に得られる森林成立初期に生まれた個体は太い幹をつくり、高さ成長にはあまり投資しない。一方、森林成立後のやや暗い環境で生まれた個体は、初め高さ成長に投資して細い幹を形成するが、光環境が改善されないとすぐにそれをやめて太さ成長(光合成のための通導器官のパイプ径)への投資の割合を高める。個体の成長過程でこのような「日和見」的性質を持つことが、森林の遷移の過程でアラカシが個体群を維持できる理由の一つと考えられる。また、個々の樹木の幹への投資パターンを生育環境にからめてモデル化することにより、個体群構造や森林構造の変化を説明・予測することも可能となった。
動き回ることができる動物たちと違って、樹木の競争は隣り合う個体間で起こる。隣接する個体間の相互作用を調べるためには、葉群の支持器官である枝や幹の空間構造を把握しておくことも重要である。測定機器の発達により、最近は森林レベルの三次元構造を非破壊的に調べることが可能となった。図のように三次元構造をデジタル化することは、より高度な構造解析に不可欠である。
以上のように、幹・枝や葉群の構造は、被圧(他の樹木の葉群を覆うこと)や競争(葉を広げる空間(光)の奪い合い)等に影響することで、樹木の生残や森林全体の動態に深く関わってくる。森林は最も三次元的構造の発達する陸上植物群落であり、その構造や発達様式に関する研究は森林の発達・維持の機構を解明するうえで重要な鍵のひとつである。これまで私がかかわってきた温・暖帯の森林に比べて、今後私が研究対象とする北方林では「寒さ」と「太陽光入射角の低さ」が生理的にも生態的にも樹木の成長に対する制限要因となると考えられる。これらの物理的要因が幹や枝の構造発達パターンにどのような変化をもたらし、どのような森林の全体構造が構築され、そのことが北方林の維持や更新にどのように影響するのか、について研究を進めていきたい。

落葉広葉樹林のクリ(Castanea crenata、実線)と
それを取り囲む他の樹種(点線)の三次元構造の再現。数字の単位はm
| (1) | 特別共同研究:平成12年度の課題は「寒冷陸域における植生、水、土壌の相互作用」(平成12年度まで)、「氷晶雲の放射特性に関する研究」(平成13年度まで)、の2課題です。申請者は本研究課題に沿った分担研究課題を設定して応募願います。特別共同研究1課題の校費は1年間で200万円を上限としますが、旅費は一般共同研究と比べて特に優遇はされません。 |
| (2) | 一般共同研究:平成13年度の採択件数は、前年度からの継続課題を含み60課題程度です。平成11年度1課題あたりの経費の平均は、校費3万円、旅費12万円でした。 |
| (3) | 研究集会:原則として旅費のみの申請としますが、印刷費の申請も可能です。 |
subject: applyとし、本文は以下のようにして下さい。
本文はa.から書き始めて下さい。
a.「特別」「一般」「集会」の別
b. 申請者氏名
c. 申請者所属
d. 申請者職名
e. 低温科学研究所以外に所属する共同研究者氏名(所属、職名)
f. 低温科学研究所に所属する共同研究者氏名
g. 共同研究課題
h.「新規」「継続」の別
i. 経費の内、消耗品・設備費等の総額(要求額)(千円)
j. 経費の内、旅費の総額(要求額)(千円)
k. 旅費の総額の内、運賃額(要求額)(千円)
l. 旅費の総額の内、宿泊費(要求額)(千円)
m. 経費の合計額(千円)
注)e, fで複数の場合は;(セミコロン)で区切って下さい。
(見本)
To: kyodo@pop.lowtem...
(... は .hokudai.ac.jp と読みかえること)
subject: apply
----------------- 以 下 本 文 ----------------
a.一般
b.東海林明雄
c.北海道教育大学教育学部釧路校物理学科
d.教授
e.鷹尾元(森林総研北海道支所、研究官);町村尚(北大農、助手);矢澤健司(科技庁航空宇宙技術研、研究室長)
f.福田正己;串田圭司
g.釧路湿原からのメタンガス発生にかかわる環境要素についての研究
h.新規
i.200
j.250
k.130
l.120
m.450
| 日付 | 内容 | 氏名 | 旧職(現職) |
|---|---|---|---|
| 13.12.31 | 辞職 | 金子あかね | 第一研究協力室事務補助員 |
| 14. 1. 7 | 復職 | 松本 慎一 | 技術専門職員 |
| 14. 1.10 | 採用 | シモネイ,バーント | 外国人研究員・客員教授 |